
Luis Gómez-Orellana, Pablo Ramil-Rego, Javier Ferreiro da Costa, Castor Muñoz Sobrino
Publicado por primera vez: 15 junio 2021
Boreas. https://doi.org/10.1111/bor.12535. ISSN 0300-9483
Artículo original Acceso Abierto
https://onlinelibrary.wiley.com/share/QZSKRZSASTTZYSRYUZD6?target=10.1111/bor.12535
A formación sedimentaria e o bosque fósil de Ponzos (sociedadecolumba.com)
Introducción
El entorno intermareal de la playa de Ponzos (NW Península Ibérica) alberga una secuencia sedimentaria (incluyendo grandes fragmentos de madera) depositada durante la primera mitad del Holoceno en un humedal continental higrófilo. Los datos de polen y macrofósiles junto con la datación por radiocarbono permiten reconstruir los cambios que ocurrieron durante el Holoceno Temprano y Medio en el paisaje de las tierras bajas costeras del noroeste de Iberia, así como en las comunidades locales de plantas de humedales, en respuesta a las variaciones climáticas y las oscilaciones eustáticas del nivel del mar. La secuencia representa la evolución de un humedal costero desde sus fases iniciales como humedal higrófilo hacia la posterior instalación de una laguna de agua dulce. Los datos de polen muestran el papel dominante de los taxones leñosos atlánticos (principalmente caducifolios), la escasez de coníferas y la falta de elementos mediterráneos en los paisajes costeros alrededor del sitio de Ponzos. La presencia y abundancia de algunos taxones como Quercus caducifolio, Castanea, Fagus, Tilia y Ulmus durante el Holoceno Temprano proporciona un apoyo adicional para la aparición de refugios glaciares en el área cantábrico-atlántica durante la Última Glaciación. La diversa vegetación que caracteriza los paisajes modernos en este territorio se estableció más tarde, extendiéndose a partir de estos reservorios glaciares de biodiversidad. En este sentido, cabe destacar la notable y temprana presencia de Fagus a principios del Holoceno, árbol también registrado previamente durante varias fases del Último Ciclo Glacial en las costas del NW Iberia. Además, durante el Holoceno Temprano y Medio se registran otros árboles que actualmente están extirpados como taxones naturales en la zona, como Pinus, Tilia y Carpinus.
La información paleoecológica obtenida en Europa durante los últimos 30 años ha confirmado que ciertas zonas de las penínsulas ibérica, italiana y balcánica habrían actuado como zonas de refugio para la flora termófila y mesófila durante el Pleistoceno. Desde estas áreas, durante el Holoceno, esos taxones se extendieron y expandieron hacia otros territorios cercanos. La ubicación de las áreas de refugio para taxones termófilos y mesófilos se ha relacionado con territorios de baja altitud, ubicados por debajo de las altitudes montañosas afectadas por la nieve permanente o semipermanente. Las áreas de refugio se identifican con sistemas de valles estrechos y depresiones, donde diferentes especies podrían acomodar sus áreas de distribución de acuerdo con las fluctuaciones climáticas, y también expandirse durante los períodos de condiciones más favorables (Ramil-Rego et al. 1998a, 2000; Carrión et al. 2003; Gómez-Orellana et al. 2007, 2012, 2013; González-Sampériz et al. 2010; Tzedakis et al. 2013; Birks 2019).
En la Península Ibérica, el registro paleoecológico muestra tres patrones principales de historia vegetal que se pueden identificar, con al menos tres biorregiones. En primer lugar, la biorregión atlántica, que encierra los territorios litorales, sublitorales y montañosos a lo largo de la costa norte (litoral cantábrico) y se extiende a lo largo de la costa atlántica. Cubre áreas que durante la mayor parte del Holoceno registraron temperaturas invernales suaves y ausencia de sequías de verano. En segundo lugar, la parte sur del litoral atlántico y su territorio análogo en la costa mediterránea perteneciente a la biorregión mediterránea, con un clima durante el Holoceno marcado por inviernos suaves y veranos cálidos y secos. En tercer lugar, el interior de la Península Ibérica, formado por extensas mesetas y diversas cadenas montañosas, formando la biorregión continental ibérica que durante el Holoceno tuvo condiciones climáticas que eran cálidas y secas en verano, y muy frías en invierno.
La presencia de áreas de refugio para la flora termófila y esclerófila se ha argumentado a partir de diversas secuencias de polen obtenidas en la biorregión mediterránea (por ejemplo, Carrión et al. 2003), vinculado a la persistencia de taxones como Quercus ilex, Olea, Phillyrea, Pistacia, Rhamnus y Buxus. Asimismo, en estudios realizados en la biorregión atlántica, la evidencia palinológica atestigua la ocurrencia de áreas de refugio para diversos elementos mesófilos como Quercus caducifolio, Corylus, Tilia, Ulmus, Castanea, Fagus, Carpinus, e incluso taxones de hoja perenne como Quercus ilex, Ilex aquifolium y Laurus nobilis (Ramil-Rego et al. 1998a, 2000; Gómez-Orellana et al. 2012, 2013). El conocimiento de la biorregión continental ibérica es todavía escaso y fragmentado y no permite valorar en profundidad la posible presencia de refugios, más aún cuando las secuencias disponibles proceden de zonas montañosas situadas en la transición entre las biorregiones mediterráneas y atlánticas (por ejemplo, Morales-Molino & García-Antón 2014).
Existen cuatro unidades biogeográficas principales dentro de la biorregión atlántica. Estos han mantenido, a través del Máximo Tardiglaciar y el Holoceno, características comunes y diferenciadas en relación con las especies presentes y los paisajes dominantes (Ramil-Rego et al. 2006). La primera unidad comprende los territorios litorales, desde el nivel del mar hasta los 200 m s.n.m. (Fig. 1). La segunda unidad incluye los territorios sublitorales, lejos de la costa y extendiéndose hacia las principales unidades montañosas. La tercera unidad corresponde a la cordillera atlántica en su conjunto, con un sistema principal, la Cordillera Cantábrica (700-2650 m s.n.m.), que se extiende E-W, y otros relieves de menor entidad y altitud situados al este y al oeste de esa cordillera principal (Fig. 1). La cuarta unidad corresponde a un área de transición entre la unidad de montaña atlántica y la biorregión continental ibérica que discurre por un conjunto de cuencas hidrográficas y valles definidos por los relieves montañosos (Ramil-Rego et al. 1998a).

Ubicación del humedal de Ponzos y otras lagunas costeras actualmente existentes en la zona
(1 = Depósito de Ponzos; 2 = Laguna de Doniños; 3 = Laguna de Frouxeira; 4 = Laguna de Pantín).
Los registros de alta resolución para el Máximo Tardiglaciar y el inicio del Holoceno en la biorregión atlántica de la Península Ibérica corresponden a depósitos formados en diferentes tipos de humedales (pantanos de manta, turberas, lagos, lagunas, etc.). La mayoría de ellos se encuentran en la unidad montañosa, particularmente en la Cordillera Cantábrica y en otras montañas cercanas: la Sierra Del Norte de Galicia (Van Mourik 1986; Ramil-Rego et al. 1996, 1998a; Muñoz Sobrino et al. 2005), la Sierra Gallega-Miño (Ramil-Rego et al. 1998a) y la Sierra Gallega-Duero (Allen et al. 1996; Muñoz Sobrino et al. 2004, 2007, 2013) (Fig. 1). La mayoría de estos depósitos están lejos de las tierras bajas costeras, donde probablemente se habrían ubicado las principales áreas de refugio para los taxones mesófilos durante el Pleistoceno. Así, en la actualidad, un gran número de flora mesófila localizada en otros territorios de la región biogeográfica atlántica no están presentes en estas zonas interiores.
Los análisis paleobotánicos realizados sobre los sedimentos de los diferentes tipos de humedales litorales y sublitorales de la biorregión atlántica ibérica han proporcionado información desigual y fragmentada. La causa de esta dispersión desigual de los datos radica en las características geomorfológicas (ausencia de grandes cuencas o escenarios favorables al establecimiento de grandes humedales, efectos de los procesos erosivos, cambios en la configuración hidrológica superficial y en la línea costera, etc.). Además, la acción humana ha ido destruyendo progresivamente estos yacimientos debido al aumento de las actividades agrícolas, forestales y urbanas.
El final del periodo glacial marcó un punto de inflexión en los ecosistemas litorales de las costas cantábrica y atlántica de la Península Ibérica. El clima más cálido y húmedo favoreció la difusión y expansión de varios taxones, y el aumento del nivel del mar provocó la reconfiguración espacial de los ecosistemas. Algunas secciones litorales perdieron parte de sus hábitats costeros (dunas, humedales, acantilados), que fueron destruidos por la progresión interior de la costa, mientras que en otros tramos los hábitats costeros se reconfiguraron o incluso se establecieron en los antiguos territorios continentales. En relación con estas dinámicas, se han identificado diferentes restos limnicos como antiguos depósitos orgánicos formados en humedales continentales en el litoral. Hoy en día, estos restos se encuentran entre las áreas eulitoral e infralitoral, y registran diferentes períodos del último ciclo glacial-interglacial. Sólo la secuencia palinológica del Área Longa cubre el último período glaciar (Gómez-Orellana et al. 2007). Varios sitios de polen registran diferentes períodos del MIS 3 (Nonn 1966; Mary et al. 1977; Granja & De Groot 1996; Gómez-Orellana et al. 2007, 2013); y otros grupos de sitios registran la segunda mitad o el último tercio del Holoceno (Santos et al. 2001; García Antón et al. 2006; Bao et al. 2007; Costas et al. 2009; Granja et al. 2010, 2016; Gómez-Orellana et al. 2014; Muñoz Sobrino et al. 2016; Sáez et al. 2018). Finalmente, las secuencias de polen que registran el Lateglacial o el comienzo del Holoceno se limitan a los depósitos de Mougás (Gómez-Orellana et al. 1998) y Urdaibai (Iriarte Chiapusso et al. 2006).
La presencia de antiguos depósitos continentales de humedales que permanecen permanente o temporalmente sumergidos en el mar también se ha encontrado en diferentes áreas atlánticas, como en el caso de la zona de Doggerland (Mar del Norte). Esta área es un amplio espacio continental que sirvió como puente entre las Islas Británicas y el continente europeo, pero que se ha mantenido por debajo del nivel del mar desde 8450-8150 aC (Gaffne et al. 2007; Coles 2014; Krüger et al. 2017).
Este trabajo presenta los datos paleoecológicos obtenidos de un yacimiento fósil acumulado en un antiguo humedal situado en la actual playa de Ponzos, en el NW de la Península Ibérica (Galicia, España). El yacimiento está situado en el extremo norte del litoral atlántico, en el tramo costero designado como 'Costa Ártabra' (Fig. 1), a tan solo 50 km en línea recta de la zona con mayor altitud de la Sierra Norte de Galicia (Montañas Xistrales; Fig. 1). Esta zona de montaña proporciona una amplia gama de información paleobotánica obtenida de diferentes tipos de turberas y depósitos orgánicos ubicados entre 650 y 1050 m s.n.m. (Ramil-Rego et al. 1996, 1998a; Muñoz Sobrino et al. 2005; Iriarte Chiapusso et al. 2016). Entre esta cordillera y el litoral, la unidad sublitoral incluye valles fluviales pequeños y estrechos orientados al N-S o W-E, donde se encuentran restos de bosques antiguos. Las especies caducifolias (Quercus robur, Quercus pyrenaica, Corylus avellana, Betula alba, Acer pseudoplatanus, Fraxinus excelsior, Fraxinus angustifolia, Ulmus glabra, Frangula alnus) son dominantes en estos bosques, con una escasa representación de especies de hoja perenne (Arbutus unedo, Ilex aquifolium, Taxus baccata, Laurus nobilis ). Además, estos bosques milenarios también albergan una importante flora nemoral, incluyendo la presencia de especies de helechos consideradas como paleo-relictos y que destacan por su alto valor para la conservación de la biodiversidad. El área de distribución actual de estas especies de helechos no está incluida en la ecozona paleártica: Culcita macrocarpa, Woodwardia radicans, Hymenophyllum tunbrigense, H. wilsonii, Vandenboschia speciosa (Jermy 1984; Sermolli et al. 1988; Rita 1990; Pausas & Sáez 2000; Amigo et al. 2017). Además, muestran un claro comportamiento estenoico respecto a las demandas de sus hábitats (ausencia de heladas, alta humedad continua, baja incidencia directa de luz solar) así como una baja capacidad de dispersión.
El objetivo de este estudio es reconstruir el paisaje de las tierras bajas costeras del noroeste de Iberia durante las fases de expansión y dominación arbórea en el Holoceno, así como descifrar la dinámica de los humedales costeros en relación con la transgresión marina del Holoceno.
Material y métodos
Muestreo
El litoral de Costa Ártabra (Fig. 1) está dominado por rocas graníticas (granitos de dos micas y leucogranitos) combinadas con otros materiales geológicos. Los procesos geomorfológicos han propiciado la aparición de largos sectores de acantilados y playas de guijarros, en contraste con pequeñas bahías que tienen, en sus tramos interiores, amplias playas de arena, estuarios y diferentes tipos de humedales costeros (lagunas, marismas, y ambientes higrófilos y pantanosos). La playa de Ponzos (latitud 43°33′N, longitud 8°15′O) tiene 1,2 km de largo (Figs 1, 2). Detrás de la playa hay un amplio y complejo sistema dunar de más de 100 hectáreas, formado por dunas embrionarias, dunas blancas móviles de Ammophila arenaria y dunas grises fijas con vegetación herbácea y pequeños arbustos (Daphne, Calluna, Erica, Ulex, Helichrysum, Othantus ). La duna gris incluye pequeñas depresiones húmedas con encharcamiento y alta humedad entre otoño y primavera, disminuyendo o casi desapareciendo con la llegada del verano. La transición al medio continental se produce sobre una zona de dunas fósiles cubiertas por arbustos dominados por vaganos de Erica y pequeñas representaciones de brezales secos en las dunas, humedales higrófilos con formaciones herbáceas y boscosas, y pequeñas representaciones de ambientes rocosos (Fig. 2).
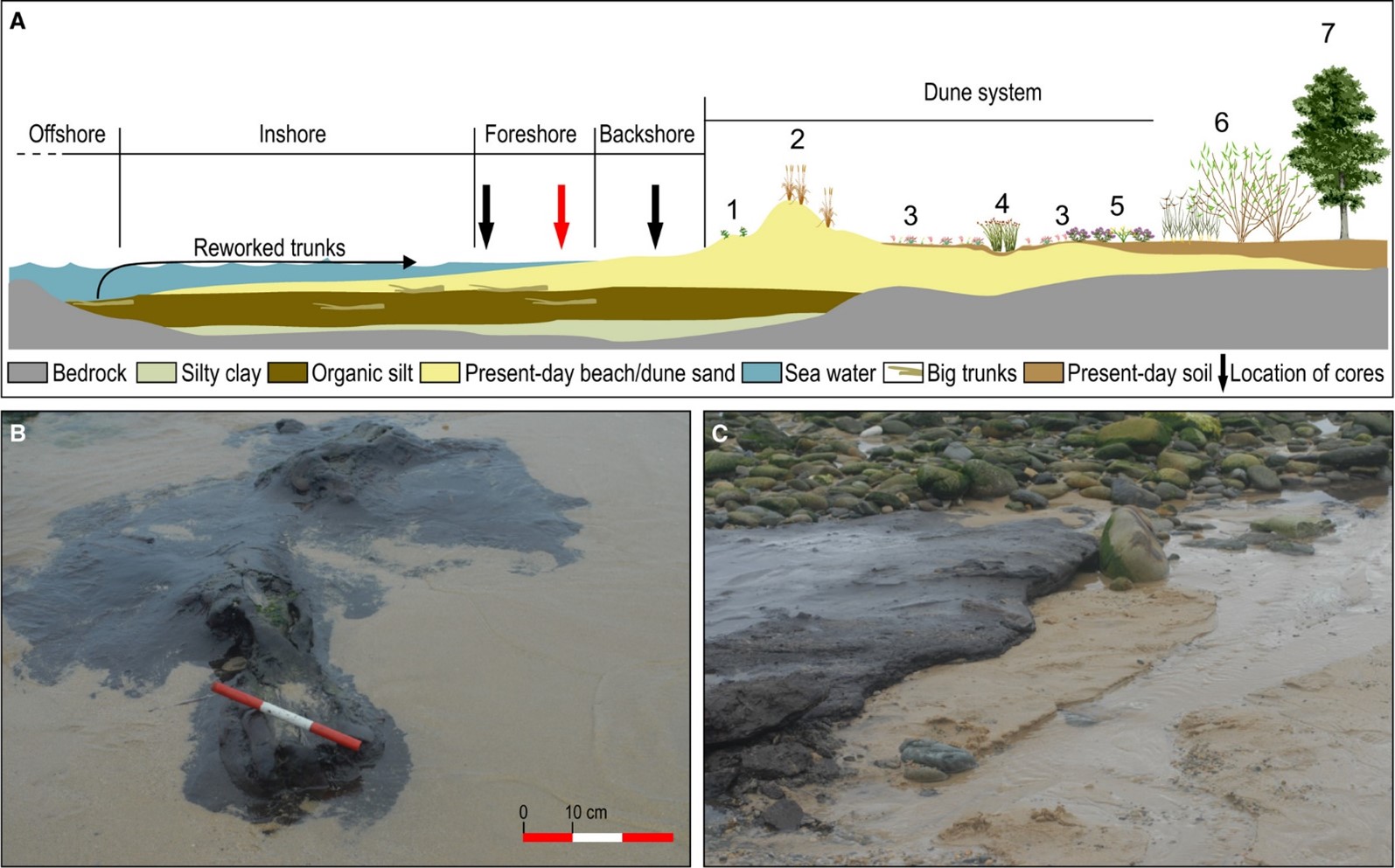
A. Sección idealizada del yacimiento de Ponzos, indicando las diferentes facies presentes, los puntos de muestreo (la flecha roja muestra la ubicación del núcleo analizado) y la configuración de la zona. 1 = dunas embrionarias cambiantes; 2 = dunas cambiantes a lo largo de la costa con Ammophila arenaria (dunas blancas); 3 = dunas costeras fijas con vegetación herbácea (dunas grises); 4 = pantalones de dunas húmedas; 5 = Dunas fijas descalcificadas del Atlántico; 6 = humedales herbáceos y leñosos; 7 = bosque. B. Fotografía de un baúl en Ponzos. C. Afloramiento de sedimento de turba en Ponzos.
Durante los períodos de mareas vivas, un grueso paquete de sedimentos orgánicos aparece en el borde occidental de la playa, en el área intermareal. Este depósito normalmente permanece cubierto por la arena de la playa actual. Este sedimento orgánico fue monitoreado continuamente de 2009 a 2020 para aprovechar las mareas vivas con el fin de documentar y recolectar diferentes macro-restos. La superficie de afloramiento del paquete de sedimentos orgánicos alcanza los 900 m2, que comprende un gran número de fragmentos de madera, entre los que destaca la aparición de troncos o ramas largas (>5 m de largo y >40 cm de diámetro). Se ubican por encima del depósito o embebidos, y se distribuyen caóticamente alrededor de diferentes áreas del afloramiento. Cuando el sedimento que rodea los restos de madera se eliminó manualmente, se determinó que algunos de estos están en posición de vida e incluyen tanto las raíces principales como la parte inicial del tronco. Durante el muestreo, algunos de estos macro-restos fueron recolectados para la identificación botánica y la datación.
Perforamos la superficie del depósito en diferentes sitios con equipos de perforación por percusión Eijkelkamp, utilizando núcleos de 10 y 3 cm de diámetro. Todas las sondas llegaron al sótano rocoso. Los núcleos de perforación recuperados en las diferentes pruebas fueron muy similares. Las principales diferencias se refieren al grosor del paquete orgánico y las características de los niveles superiores (ausencia o presencia de arenas y grandes restos de madera). El núcleo más grueso (250 cm) obtenido fue elegido para el análisis palinológico y cronológico. La litología del núcleo se describió en el laboratorio y luego se dividió en 125 muestras de 2 cm de espesor. Todas las muestras permanecieron almacenadas a 5 °C hasta el análisis.
Análisis de polen
Las muestras se prepararon para el análisis de polen utilizando métodos estándar (Fægri et al. 1989; Moore et al. 1991). Las diapositivas montadas se analizaron mediante microscopía de luz. La suma total de polen en todas las muestras oscila entre 411 y 789 granos. Los porcentajes de polen terrestre se calcularon utilizando una suma total de polen terrestre (>300 granos), incluidos árboles, arbustos y hierbas de las tierras altas. La suma total de polen se utilizó para calcular porcentajes de los acuáticos y criptogramas. El software TILIA 1.7.14 (Grimm 1990-2015) se utilizó para los cálculos y la representación gráfica de los resultados. Los diagramas de polen se zonificaron mediante el análisis de conglomerados de suma incremental restringida de cuadrados (CONISS) (Fig. 3).

Diagrama de porcentajes de polen de los taxones arbóreos registrados en el depósito de Ponzos. Biozonas: He = Expansión arbórea del Holoceno; Hd = Holoceno medio, de Muñoz Sobrino et al. El modelo de profundidad/edad se da a la izquierda, basado en la datación y el evento regional palinoestratigráfico bien fechado, relacionado con la expansión de Corylus (8500 cal. a BP).
Cronología
Se obtuvieron cinco fechas de radiocarbono para el núcleo, cuatro utilizando métodos de datación AMS y uno más utilizando los convencionales Método 14C (Tabla 1). Las determinaciones de la edad 14C se llevaron a cabo en el Centrum voor Isotopen Onderzoek, Universidad de Groningen y en el Laboratorio ICA, Florida. Las edades de radiocarbono se convirtieron en cal. a BP (2σ) usando Calib 8.2 (Stuiver et al. 2020; Tabla 1).
|
Laboratorio no. |
Profundidad (cm) |
Material |
Edad de radiocarbono uncal. a BP |
Método |
2σ cal. a Rangos de años BP (área relativa) |
Cal. de la mediana para los años BP |
|---|---|---|---|---|---|---|
|
GrN 32293 |
0 |
Salix |
6.075 ± 35 |
Convencional |
7.149–7.126 (0.03) 6.841–7.017 (0.94) 6.838–6.833 (0.00) 6.818–6.798 (0.03) |
6.929 |
|
ICA-0OS/0901 |
20–22 |
Sedimento |
5.380 ± 40 |
AMS |
6.171–6.283 (0.65) 6.103–6.158 (0.18) 6.010–6.081 (0.16) |
6.227 |
|
GrA-65937 |
52–54 |
Sedimento |
5.775 ± 40 |
AMS |
6.479–6.668 (1) |
6.573 |
|
GrA 66804 |
56 |
Salix |
5.860 ± 40 |
AMS |
6.778–6.763 (0.03) 6.596–6.756 (0.91) 6.594–6.562 (0.06) |
6.676 |
|
GrA-65936 |
140–142 |
Sedimento |
6.720 ± 40 |
AMS |
7.555–7.663 (0.85) 7.510–7.544 (0.15) |
7.609 |
Tabla 1. Radiocarbono y edades calibradas de Ponzos. Todas las fechas se calibraron utilizando el programa CALIB Rev 8.2 y el conjunto de datos IntCal20 (Stuiver et al. 2020). Se utilizaron los intervalos de confianza de 2-sigma (95,4%) y sus áreas relativas, así como el cálculo de la mediana para los años BP.
Resultados
Bioestratigrafía y fechas de radiocarbono
Los núcleos obtenidos incluyen diferentes facies orgánicas e inorgánicas, sus bases se colocan directamente sobre el lecho rocoso. El núcleo más profundo obtenido en la playa de Ponzos alcanzó una profundidad de 250 cm y comprende varias facies bien diferenciadas (Fig. 3). La parte basal (250-154 cm de profundidad) está formada por arcilla limosa gris-marrón, con diferentes tonos marrones, probablemente relacionados con el cambio en el contenido de materia orgánica. El tono marrón aumenta de abajo hacia arriba y es posible distinguir hasta tres facies (Figs. 2, 3). Entre 216 y 236 cm de profundidad, hay un alto contenido de arenas heterométricas de cuarzo. Arriba, hay una capa limosa muy oscura con alto contenido de materia orgánica y presencia de restos vegetales. Tiene un grosor de 154 cm, con grandes fragmentos de madera incrustada de Salix (Figs. 2, 3). En la parte superior de esta capa, hay un conjunto importante de macro-restos de plantas de gran tamaño (Figs. 2, 3). El nivel superior es un paquete de arena, de 75 a 100 cm de espesor, correspondiente a la playa actual.
Se obtuvieron cinco dataciones por radiocarbono, tres a partir de sedimentos a granel y dos de dos fragmentos de madera (Tabla 1). Uno de los fragmentos fue recuperado a 56 cm de profundidad desde el interior del núcleo muestreado, incrustado dentro del sedimento. El otro fue recuperado dentro de los macro-restos que aparecen en la superficie del sedimento.
Análisis paleobotánicos
Los sedimentos inorgánicos y orgánicos de los humedales halófilos activos (ambientes infralitorales y eulitorales de marismas y lagunas costeras) ubicados en la biorregión atlántica se caracterizan por un alto contenido en sal, y forman eflorescencias cuando están secos. Estos suelen tener macro-restos de conchas, caparazones, espículas u otras estructuras carbonatadas pertenecientes a invertebrados marinos, así como micro-restos característicos de biocenosis halófita. No aparece ninguna eflorescencia en los sedimentos secos obtenidos en Ponzos. Además, el análisis de los residuos de 10× y 30× aumento después de tamizar 0,5, 0,1 y 0,05 mm no encontró evidencia de restos de invertebrados marinos, ni se observó ninguna reacción carbónica después del ataque con ácido. La totalidad de los macro-restos recuperados corresponde a plantas vasculares, restos fibrosos, raíces pequeñas (<2 mm de diámetro), así como trozos de hojas y tallos de Poaceae y Cyperaceae.
Las diferentes facies incluyen abundantes macro-restos vegetales, especialmente en el nivel más orgánico (0-154 cm). La mayoría de ellos corresponden a tejidos fibrosos de plantas vasculares herbáceas y fragmentos leñosos de diferentes tamaños. Los restos leñosos más grandes corresponden a 25 ramas grandes y troncos de más de 2 m de largo (hasta 5–7 m) y 30–40 cm de diámetro. Algunos de estos grandes fragmentos corresponden a raíces y fragmentos basales del tronco en disposición viva. Sin embargo, la mayoría de ellos se encuentran en la superficie del depósito, ya sea incrustados en él o libres. La mayoría de nuestras muestras leñosas son de sauce (según la flora regional actual, muy probablemente corresponden a Salix atrocinerea) y, en menor medida, de robles caducifolios (según la flora regional actual, muy probablemente corresponden a Quercus robur/Quercus pyrenaica).
El contenido de micro-restos consiste en abundante polen, así como esporas de musgo y helecho, en buen estado de conservación. Además, se encontraron varios restos de hongos (esporas, hifas), así como diatomeas y otros microfósiles no polínicos. El contenido en micro-restos es similar a la señal de polen obtenida en diferentes humedales no halófilos de la zona litoral (turberas, brezales húmedos, marismas altas, zonas pantanosas que rodean lagunas costeras, etc.).
Las figuras 3, 4 y 5 muestran los diagramas de polen obtenidos. Las zonas de polen identificadas siguen los resultados del análisis por conglomerados. Diez zonas locales de ensamblaje de polen (LPAZ) fueron reconocidas y designadas como Zonas Ponzos (PZ), pero algunas de ellas se subdividieron para resaltar algunos hechos particulares relacionados con la dinámica de la vegetación.

Diagrama de porcentajes de polen de las hierbas y arbustos registrados en el depósito de Ponzos. Biozonas: He = Expansión arbórea del Holoceno; Hd = Holoceno medio, de Muñoz Sobrino et al.

Diagrama de porcentajes de polen de los taxones vinculados a ambientes de agua dulce registrados en el depósito de Ponzos. Biozonas: He = Expansión arbórea del Holoceno; Hd = Holoceno medio, de Muñoz Sobrino et al.
PZ-1 (250–248 cm) que marca el inicio de la secuencia se caracteriza por la dominancia de los árboles, incluyendo un máximo de Quercus robur-type y alrededor del 20% de Corylus.
PZ-2 (248-238 cm) comienza con una ligera disminución en el polen total de los árboles, marcada por la disminución del tipo Quercus robur, pero la posterior expansión y máxima de Corylus.
PZ-3 (238-192 cm) evidencia la recuperación del polen total de los árboles (>85%). Las especies de árboles caducifolios dominantes son Quercus robur-type y Corylus, con escasa presencia de Pinus. En la subzona 3a hay un ligero aumento de Poaceae mientras que en la subzona 3b Erica disminuye y Compositae aumenta.
PZ-4 (192-140 cm) refleja un ligero detrimento total del polen de los árboles debido a la disminución de Corylus. Fagus y Arbutus aumentan y comienza la curva continua de Ulmus. Los porcentajes de Poaceae aumentan y los porcentajes de Erica y Compositae disminuyen. La parte superior de la zona ha sido fechada en 7.609 cal. a BP (Tabla 1, Fig. 3).
PZ-5 (140–134 cm) refleja un detrimento en el polen total de los árboles y representa el mínimo arbóreo de la secuencia, junto con el máximo de Poaceae (Fig. 4).
En PZ-6 (134-120 cm) hay un aumento de los porcentajes de polen de los árboles que refleja la recuperación de los bosques caducifolios, con la expansión sucesiva de Quercus robur-type y Corylus, y Arbutus y Fagus aumentando. Además, se produce un ligero aumento de Erica, mientras que las Poaceae disminuyen (Fig. 4).
PZ-7 (120–44 cm) representa una disminución inicial del polen de los árboles (PZ-7a) y su posterior recuperación (PZ-7b). La subzona PZ-7c refleja un aumento del tipo Quercus robur, seguido de aumentos de Corylus y Betula y los valores mínimos de Arbutus (PZ-7d). La subzona PZ-7e muestra un nuevo aumento en Quercus robur-type, Erica y Poaceae (Figs. 3, 4). La parte superior de la subzona PZ-7e ha sido fechada en 6573 cal. a BP (Tabla 1, Fig. 3). Finalmente, PZ-7f muestra la sustitución de Corylus por Quercus robur-type (Fig. 3) y el aumento continuo de Poaceae (Fig. 4).
PZ-8 (44-26 cm) muestra un momento de detrimento arbóreo, debido principalmente a disminuciones en Corylus y Quercus robur-type.
PZ-9 (26-10 cm) refleja una recuperación posterior del polen total de los árboles que se inicia con el aumento de Betula y Corylus (PZ-9a) y luego quercus robur-type (PZ-9b). La subzona PZ-9a ha sido fechada en 6227 cal. a BP (Tabla 1, Fig. 3). Quercus robur-type y Corylus disminuyen en PZ-10 (10-0 cm), mientras que Betula, Fagus, Pinus pinaster-type y Erica aumentan.
Los niveles más bajos de la secuencia no pudieron ser datados por técnicas de radiocarbono debido a su escaso contenido de materia orgánica. Por encima de ellos, se obtuvieron diferentes fechas a partir de sedimentos a granel y macroreservidos (Tabla 1, Fig. 3).
La interpretación y correlación de estas zonas polenales con las secuencias regionales de referencia para la ecorregión atlántica (Ramil-Rego 1992; Muñoz Sobrino et al. 2005) y el Las fechas 14C obtenidas permiten la atribución de toda la secuencia al comienzo del Holoceno. La base de la secuencia representa una fase de expansión de Corylus y detrimento tipo Quercus robur fechada antes de 7.069 cal. a BP. Esto permite estimar la edad de la base de la secuencia en algún momento entre las expansiones regionales de Quercus (10.950 cal. a BP) y Corylus (8.500 cal. a BP). La fecha de 6.227 cal. a BP (Tabla 1) obtenida en la parte superior de la secuencia (20 cm de profundidad) situaría el final de la secuencia alrededor de 6.000 cal. a BP (Fig. 3).
Continuar leyendo el artículo: Cambio ambiental del Holoceno en la costa atlántica del noroeste de Iberia como se infiere de la secuencia de humedales de Ponzos - (II)
Artículos relacionados:
A formación sedimentaria e o bosque fósil de Ponzos
Reflexiones prehistóricas sobre la arena de Ponzos
Restos de un bosque fósil en la playa de Ponzos (I) - Galería fotográfica (sociedadecolumba.com)
Restos de un bosque fósil en la playa de Ponzos (II) - Galería fotográfica (sociedadecolumba.com)
VIDEO Restos de un bosque fósil en la playa de Ponzos
Límnico
Termino descriptivo aplicado al ambiente deposicional de lagos de agua dulce. Utilizado especialmente para cuencas hulleras, en oposición a Parálico.
- Más información en: Límnico (Geología) © https://glosarios.servidor-alicante.com/geologia/limnico
https://www.igme.es/museo/fonelas/paginas/001-005-003.htm
Parálico
Término aplicado a las cuencas sedimentarias formadas en medios marinos marginales; p. ej., lagunas; cuencas litorales, etc. (Cf. Límnico).
- Más información en: Parálico (Geología) © https://glosarios.servidor-alicante.com/geologia/paralico
Eulitoral
Zona comprendida entre el límite máximo de la marea y el mínimo. También llamada intermareal.
Zonas de un ecosistema lacustre
Infralitoral
Dícese de la zona del fondo marino inmediatamente por debajo del nivel del agua y que, por tanto, permanece casi todo el tiempo sumergida y muy raramente emergida. Por encima de la zona infralitoral aparece la zona denominada eulitoral/mesolitoral, o intermareal, que está sumergida o emergida en función de las mareas. El límite inferior lo marca la máxima profundidad a la que existen fanerógamas marinas o algas fotófilas. Por debajo de la zona infralitoral, aparece la zona o piso circalitoral.


